幽门螺杆菌基因分型与结肠息肉
幽门螺杆菌基因分型与结肠息肉的研究进展
## 幽门螺杆菌基因分型与结肠息肉的研究进展
### 研究现状概述
目前检索到的文献中,**缺乏直接探讨幽门螺杆菌(*Helicobacter pylori*, Hp)基因分型与结肠息肉(colonic polyps)之间关联的系统性研究证据**。现有文献主要聚焦于Hp感染与上消化道疾病(慢性胃炎、消化性溃疡、胃癌、胃MALT淋巴瘤)的关系,以及Hp对食管腺癌可能的保护性作用[4]。关于Hp基因分型(尤其是*cagA*、*vacA*等毒力基因)与结肠息肉发生风险之间的关联,检索到的资料未提供直接数据。
### 幽门螺杆菌基因分型与致病性
Hp的致病性与其毒力基因密切相关,主要毒力因子包括:
- **细胞毒素相关基因A(*cagA*)**:仅存在于部分临床菌株的*cag*致病岛(*cag* PAI)中,通过Ⅳ型分泌系统(T4SS)转运至胃黏膜上皮细胞,引发一系列细胞因子反应。*cagA*阳性菌株感染显著增加胃癌发病风险,被认为是Hp致癌的主要毒力基因[1]。
- **空泡细胞毒素基因A(*vacA*)**:所有Hp菌株均携带*vacA*基因,但并非所有菌株均产生功能性VacA毒素。基因序列具有高度多态性,其变异与所致疾病的严重程度相关[1]。
- **宿主遗传因素**:IL-1、IL-2、IL-6、IL-8、TLR1/TLR10等基因多态性可影响Hp感染后的炎症反应强度及胃癌风险[3]。
### Hp感染与结直肠疾病的潜在关联
检索到的文献中,Hp感染与结直肠疾病的关系证据有限:
1. **Hp与结直肠癌的间接关联**:部分研究提示Hp感染可能通过影响胃酸分泌、肠道菌群组成及全身免疫炎症状态,间接影响结直肠肿瘤的发生风险,但因果关系尚未确立。
2. **Hp与缺血性心脏病/脑血管病的争议性关联**:文献指出,Hp定植与缺血性心脏病及脑血管病的关联在控制混杂因素后显著减弱,多数权威观点认为这些关联为非因果性[4]。这一方法论提示,Hp与结肠息肉之间的关联同样需警惕混杂偏倚。
### 研究空白与未来方向
1. **缺乏直接证据**:检索到的文献未涉及Hp基因分型(*cagA*+/−、*vacA* s1/m1等亚型)与结肠息肉(腺瘤性、锯齿状、增生性等)组织学类型或数量之间的关联研究。
2. **潜在机制假设**:Hp感染诱导的慢性低度炎症状态、胃泌素水平改变、肠道微生态失调等可能是连接Hp与结直肠肿瘤的潜在桥梁,但需进一步验证。
3. **流行病学挑战**:Hp感染与结肠息肉的关联研究需严格控制年龄、性别、饮食因素(高盐、腌制食品摄入)、非甾体抗炎药使用、吸烟等混杂因素[2][3]。
### 结论
现有检索证据不支持对幽门螺杆菌基因分型与结肠息肉之间的关联做出任何结论。该领域属于研究空白,需开展设计严谨的流行病学研究(如大样本病例对照研究或前瞻性队列研究),同时进行Hp毒力基因分型(*cagA*、*vacA*亚型)及结肠息肉组织学分型,以明确二者之间是否存在独立关联。
> 本内容基于检索到的文献生成,仅供临床参考。具体诊疗决策应结合患者个体情况并核实原始文献。
幽门螺杆菌 cagA vacA 基因分型 结直肠腺瘤 病例对照研究 临床证据
## 幽门螺杆菌 *cagA*/*vacA* 基因分型与结直肠腺瘤的病例对照研究:临床证据综述
### 结论
**检索到的文献中,缺乏直接探讨幽门螺杆菌(*H. pylori*, Hp)*cagA*/*vacA* 基因分型与结直肠腺瘤(colorectal adenoma)之间关联的病例对照研究或临床证据。** 现有证据主要集中于Hp毒力基因与胃癌风险的关系,以及Hp感染与上消化道疾病的关联,结直肠腺瘤方向属于明确的研究空白。
---
### 现有证据基础
#### 1. Hp毒力基因分型与致病性(已确立)
检索到的文献明确阐述了Hp主要毒力基因的生物学功能及临床意义:
| 毒力基因 | 特征 | 临床意义 |
|---------|------|---------|
| ***cagA***(细胞毒素相关基因A) | 仅存在于部分菌株的*cag*致病岛(*cag* PAI),通过Ⅳ型分泌系统(T4SS)转运CagA毒素至胃黏膜细胞 | 被认为是Hp致癌的主要毒力基因;*cagA*阳性菌株感染显著增加胃癌发病风险[1] |
| ***vacA***(空泡细胞毒素基因A) | 所有Hp菌株均携带,但非所有菌株产生功能性VacA毒素;基因序列高度多态性 | 基因变异与疾病严重程度相关;VacA毒素导致细胞质空泡化改变[1] |
| **血清学分型** | CagA/VacA抗体任一项阳性→Ⅰ型菌株(毒力较强);仅UreA/UreB阳性→Ⅱ型菌株(毒力较弱)[2] | 用于临床区分高毒力与低毒力菌株感染 |
#### 2. Hp毒力基因与胃癌风险(已确立)
- **宿主-病原体交互作用**:携带高表达IL-1β基因多态性的个体,若同时感染*cagA*+或*vacA* s1型菌株,胃癌风险分别增加**25倍**或**87倍**(相较于未感染者)[4]。
- **分子流行病学**:多数病例对照研究显示,胃癌/消化性溃疡患者中*cagA*阳性Hp感染比例高于健康对照[5]。
- **局限性**:严格来说,将分析限制于Hp阳性人群时,数据同样支持"*cagA*阴性菌株降低风险"的假设,需纳入Hp阴性对照才能完整评估[5]。
#### 3. Hp感染与结直肠腺瘤——证据缺口
检索到的文献中,**无任何内容涉及Hp感染(无论是否分型)与结直肠腺瘤的关联**。现有文献覆盖范围:
- Hp与慢性胃炎、消化性溃疡、胃癌、胃MALT淋巴瘤[1]
- Hp血清学分型检测方法[2]
- Hp根除后随访策略[3]
- 宿主遗传因素与胃癌风险[4]
- 病例对照研究的方法学讨论(以Hp*cagA*与胃癌为例)[5]
---
### 研究空白分析
| 维度 | 现状 |
|------|------|
| **直接证据** | 检索到的文献中无Hp基因分型与结直肠腺瘤的病例对照研究 |
| **间接机制假设** | Hp感染可能通过胃酸分泌改变、肠道菌群失调、全身慢性炎症等途径影响结直肠肿瘤发生,但缺乏Hp毒力基因分型层面的数据 |
| **方法学挑战** | 需控制年龄、性别、饮食(高盐、腌制食品)、NSAIDs使用、吸烟、肥胖等混杂因素;需纳入Hp阴性对照以完整评估效应方向[5] |
| **研究设计建议** | 建议开展**病例对照研究**:病例组为经结肠镜确诊的结直肠腺瘤患者,对照组为结肠镜阴性人群;对Hp阳性者进行*cagA*/*vacA*血清学分型或基因分型;同时采集Hp阴性对照 |
---
### 结论
现有检索证据不支持对幽门螺杆菌 *cagA*/*vacA* 基因分型与结直肠腺瘤之间的关联做出任何临床结论。该领域属于明确的研究空白,需开展设计严谨的病例对照研究,同时进行Hp毒力基因分型及结直肠腺瘤组织学分型,以明确二者之间是否存在独立关联。
> 本内容基于检索到的文献生成,仅供临床参考。具体诊疗决策应结合患者个体情况并核实原始文献。
幽门螺杆菌 毒力因子 结直肠肿瘤 流行病学 风险因素 混杂因素 控制
## 幽门螺杆菌毒力因子与结直肠肿瘤:流行病学证据、风险因素与混杂控制
### 结论
**检索到的文献中,无直接证据支持幽门螺杆菌(*H. pylori*, Hp)毒力因子(*cagA*/*vacA*)与结直肠肿瘤(colorectal neoplasia)之间存在独立流行病学关联。** 现有证据明确确立了Hp毒力因子与胃癌的因果关系,但结直肠肿瘤方向属于研究空白。若开展相关研究,需严格控制的混杂因素包括年龄、社会经济水平、饮食因素(高盐、腌制食品摄入)、吸烟及宿主遗传多态性。
---
### 一、已确立的Hp毒力因子与胃癌的流行病学证据
#### 1. 毒力因子的生物学基础
| 毒力因子 | 特征 | 致癌机制 |
|---------|------|---------|
| ***cagA***(细胞毒素相关基因A) | 仅存在于部分菌株的*cag*致病岛(*cag* PAI),通过Ⅳ型分泌系统(T4SS)注入宿主细胞 | CagA被宿主细胞磷酸化后,诱导上皮极性改变、增殖和迁移异常,增加癌症风险[1][3] |
| ***vacA***(空泡细胞毒素基因A) | 所有菌株均携带,但基因序列高度多态性;功能性VacA毒素导致细胞质空泡化 | 基因变异与疾病严重程度相关[1] |
| **其他毒力因子** | 可导致宿主DNA双链断裂、干扰干细胞特性、阻断感染细胞对炎症刺激的反应 | 通过多种分子通路促进癌变[3] |
#### 2. 流行病学证据
- **WHO分类**:Hp被世界卫生组织/国际癌症研究机构(WHO/IARC)定为**Ⅰ类胃癌致病因子**[1][3]。
- **终生风险**:Hp感染者终生患胃癌风险约为**1.5-2.0%**,但由于全球感染基数庞大,每年胃癌负担超过100万例[2]。
- **地域分布**:东亚(日本、韩国、中国东部)患病率最高,全球40%的胃癌病例发生在中国[2]。
- **毒力因子分层**:分子流行病学调查显示,*cagA*阳性菌株感染显著增加胃癌发病风险[1];宿主-病原体交互作用(如IL-1β基因多态性与*cagA*+或*vacA* s1型菌株的协同作用)可进一步放大风险[2]。
---
### 二、Hp毒力因子与结直肠肿瘤——研究空白
检索到的文献中,**无任何内容涉及Hp毒力因子(*cagA*/*vacA*)与结直肠肿瘤(包括结直肠腺瘤、结直肠癌)的关联**。现有文献覆盖范围严格限定于:
- Hp与慢性胃炎、消化性溃疡、胃癌、胃MALT淋巴瘤[1]
- Hp感染流行率的决定因素(社会经济水平、年龄、队列效应)[2]
- Hp致癌的分子机制(CagA/T4SS、DNA损伤、干细胞扩增)[3]
- 病例对照研究中混杂因素控制的方法学讨论[4]
**结论**:Hp毒力因子与结直肠肿瘤之间的关联属于明确的研究空白,尚无流行病学数据支持或否定这一假设。
---
### 三、若开展相关研究需控制的混杂因素
基于检索到的文献,以下混杂因素在Hp感染与结直肠肿瘤的关联研究中必须严格控制:
#### 1. 社会人口学因素
| 混杂因素 | 控制理由 | 来源 |
|---------|---------|------|
| **年龄** | Hp感染流行率随年龄增长而增加(主要为队列效应);结直肠肿瘤发病率亦随年龄增长 | [2] |
| **社会经济水平** | 儿童时期的社会经济水平是Hp感染流行率的主要决定因素;社会经济水平亦与结直肠癌筛查率、饮食模式相关 | [2] |
| **种族/民族** | 种族差异可能反映社会经济水平或文化实践差异,而非遗传基础 | [2] |
#### 2. 饮食与生活方式因素
| 混杂因素 | 控制理由 | 来源 |
|---------|---------|------|
| **高盐摄入** | 与胃癌风险增加相关;亦可能通过影响胃酸分泌间接影响肠道菌群 | [2] |
| **腌制食品摄入** | 与胃癌风险增加相关 | [2] |
| **抗氧化剂含量低的饮食** | 与胃癌风险增加相关;亦为结直肠癌风险因素 | [2] |
| **吸烟** | 宿主方面与胃癌风险增加相关的因素之一;亦为结直肠癌明确风险因素 | [2] |
#### 3. 宿主遗传因素
| 混杂因素 | 控制理由 | 来源 |
|---------|---------|------|
| **IL-1基因多态性** | 决定促炎细胞因子表达,与Hp感染后胃癌风险增加相关 | [2] |
| **病原体识别受体基因多态性** | 影响宿主对Hp感染的免疫应答强度 | [2] |
#### 4. 方法学注意事项(基于病例对照研究设计)
检索到的文献[4]指出,病例对照研究中混杂因素的控制需特别注意:
- **混杂的定义**:当某个因素与暴露(Hp感染/毒力因子)和疾病(结直肠肿瘤)均存在关联时,可产生混杂偏倚。
- **病例对照研究的特殊问题**:若病例组和对照组的招募比例在不同分层(如研究地点)中不一致,即使该因素本身与疾病风险无关,仍可产生混杂。例如,若不同地区Hp感染率与结直肠肿瘤发病率存在地域差异,且病例和对照的招募比例在各地区不均衡,则需进行分层调整或标准化[4]。
- **控制方法**:可采用分层分析、多变量回归模型(logistic回归)、倾向性评分匹配等方法。
---
### 四、研究设计建议
若拟开展Hp毒力因子与结直肠肿瘤的病例对照研究,建议:
1. **病例组**:经结肠镜确诊的结直肠腺瘤或结直肠癌患者
2. **对照组**:结肠镜阴性人群(需匹配年龄、性别、社会经济水平)
3. **暴露评估**:Hp血清学检测(区分Ⅰ型[*cagA*/*vacA*阳性]与Ⅱ型菌株)或胃黏膜活检基因分型
4. **需采集的协变量**:年龄、性别、社会经济水平、吸烟史、饮食模式(高盐/腌制食品摄入)、NSAIDs使用史、结直肠癌家族史、宿主IL-1β基因多态性
5. **分析策略**:多变量logistic回归,报告调整后的OR值及95% CI;进行亚组分析(按Hp毒力因子分层)
---
### 五、结论
现有检索证据不支持对幽门螺杆菌毒力因子与结直肠肿瘤之间的关联做出任何临床结论。该领域属于明确的研究空白。若开展相关流行病学研究,需严格控制的混杂因素包括年龄、社会经济水平、饮食因素(高盐、腌制食品摄入)、吸烟及宿主遗传多态性,并注意病例对照研究中因招募比例不均衡导致的混杂偏倚。
> 本内容基于检索到的文献生成,仅供临床参考。具体诊疗决策应结合患者个体情况并核实原始文献。
幽门螺杆菌 感染 结直肠肿瘤 风险比 荟萃分析 2023
⚠️提示:当前回答基于第三方数据源补充召回的文献,请注意甄别信息来源的可靠性。
## 幽门螺杆菌感染与结直肠肿瘤风险:2023年荟萃分析证据综述
### 结论
**现有高质量荟萃分析一致表明,幽门螺杆菌(*H. pylori*, Hp)感染与结直肠肿瘤(包括结直肠腺瘤、进展期腺瘤及结直肠癌)风险增加存在统计学显著的正向关联,但基于观察性研究的设计,因果关系尚未确立。**
---
### 一、核心荟萃分析证据汇总
#### 1. 2025年大规模荟萃分析(Li X et al.)
| 结局指标 | OR (95% CI) | I² | 纳入研究数 | 总样本量 |
|---------|------------|-----|-----------|---------|
| 结直肠癌 | 1.59 (1.39–1.82) | 89% | 55项 | 48,945,236 |
| 结直肠腺瘤 | 1.47 (1.36–1.59) | 88% | 同上 | 同上 |
| 进展期腺瘤 | 1.77 (1.56–2.00) | 65% | 同上 | 同上 |
| 总体结直肠肿瘤 | 1.49 (1.39–1.60) | 91% | 同上 | 同上 |
该研究同时纳入一项中国浙江筛查队列的原始数据(n=3,475),结果显示Hp感染与结直肠癌(OR=2.57, 95% CI: 1.26–5.39)及腺瘤(OR=1.37, 95% CI: 1.19–1.59)均显著相关,其中进展期腺瘤的关联强度更高(OR=1.94, 95% CI: 1.59–2.38)[2]。
#### 2. 2025年系统综述与荟萃分析(Kim JH et al.)
纳入40项研究、共503,365名参与者,结果显示:
| 结局指标 | OR (95% CI) |
|---------|------------|
| 良性结直肠息肉(BCP) | 1.43 (1.29–1.58) |
| 结直肠腺瘤(CRA)及进展期CRA | 1.71 (1.41–2.08) |
**亚组分析**显示显著的种族差异:亚洲人群OR为1.99 (95% CI: 1.42–2.80),西方人群OR为1.37 (95% CI: 1.22–1.54),组间差异有统计学意义(P=0.04)[4]。
#### 3. 2020年荟萃分析(Choi DS et al.)
纳入48项研究、171,045例患者,结果:
| 结局指标 | 合并OR (95% CI) |
|---------|----------------|
| 结直肠腺瘤 | 1.49 (1.37–1.62) |
| 进展期腺瘤 | 1.50 (1.28–1.75) |
| 结直肠癌 | 1.44 (1.25–1.71) |
校正发表偏倚后,腺瘤风险仍显著增加(OR=1.39)[14][19]。
#### 4. 2019年荟萃分析(Yang F et al.)
纳入27项研究、14,357例病例,显示Hp感染与结直肠癌风险增加相关(OR=1.34, 95% CI: 1.14–1.57, P<0.001),亚组分析显示血清学检测亚组OR=1.20 (95% CI: 1.08–1.34),多种检测方法亚组OR=2.63 (95% CI: 1.09–6.31)[11]。
---
### 二、Hp毒力因子与结直肠肿瘤风险
检索到的文献中,关于Hp毒力因子(*cagA*/*vacA*)与结直肠肿瘤的直接证据有限:
- **Butt J et al.(2019)**:一项纳入美国多个前瞻性队列的大型研究显示,Hp VacA特异性血清阳性与结直肠癌风险增加相关(OR=1.11, 95% CI: 1.01–1.22)[7][8]。
- **2013年荟萃分析**:明确指出该研究"统计效能不足以确定细胞毒素相关基因A(*cagA*)阳性Hp与结直肠肿瘤风险的关系"[16]。
---
### 三、Hp根除治疗对结直肠肿瘤风险的影响
基于3项回顾性研究的荟萃分析显示,抗Hp治疗与结直肠肿瘤总体风险降低相关(OR=0.44, 95% CI: 0.20–0.97),但异质性较高(I²=93%),需谨慎解读[2]。
---
### 四、潜在机制
检索到的文献提及以下可能机制[19][23]:
1. **高胃泌素血症(Hypergastrinemia)**:Hp感染导致胃酸分泌改变,刺激胃泌素释放,胃泌素具有促进结直肠黏膜增殖的作用。
2. **肠道菌群失调(Intestinal Dysbiosis)**:Hp感染可诱导肠道促炎反应,并促进黏液降解菌群(mucus-degrading microbiota)的增殖,从而驱动结直肠癌变[23]。
3. **全身慢性炎症**:Hp感染诱导的全身性低度炎症状态可能促进结直肠肿瘤发生。
---
### 五、研究局限性与混杂因素
| 局限性 | 说明 |
|--------|------|
| **观察性研究设计** | 所有荟萃分析均基于观察性研究,无法确立因果关系[14] |
| **高异质性** | 多数荟萃分析I²>80%,提示研究间存在显著异质性 |
| **混杂因素控制不足** | 年龄、社会经济水平、饮食因素(高盐/腌制食品)、吸烟、PPI使用等混杂因素在各原始研究中控制程度不一 |
| **检测方法差异** | 血清学检测(IgG抗体)与粪便抗原检测、呼气试验、胃镜活检等方法间存在差异[16] |
| **发表偏倚** | 部分荟萃分析检测到发表偏倚,校正后效应量有所降低[14][19] |
| **毒力因子数据缺乏** | 多数研究未区分Hp毒力基因型(*cagA*/*vacA*),无法评估不同毒力菌株的风险差异[16] |
---
### 六、结论
截至2023年,多项大规模荟萃分析一致显示幽门螺杆菌感染与结直肠腺瘤(OR约1.47–1.71)、进展期腺瘤(OR约1.50–1.77)及结直肠癌(OR约1.39–1.59)风险增加存在统计学显著的正向关联。然而,基于观察性研究的设计、高异质性及混杂因素控制不足,**该关联的因果关系尚未确立**。关于Hp毒力因子(*cagA*/*vacA*)与结直肠肿瘤风险的关系,现有证据极为有限,属于明确的研究空白。
> 本内容基于检索到的文献生成,仅供临床参考。具体诊疗决策应结合患者个体情况并核实原始文献。
幽门螺杆菌 毒力因子 萎缩性胃炎 肠化 胃癌流行病学 风险因素 混杂因素 控制
## 幽门螺杆菌毒力因子与胃癌流行病学:萎缩性胃炎/肠化生背景下的风险因素与混杂控制
### 结论
**幽门螺杆菌(*H. pylori*, Hp)被WHO/IARC列为Ⅰ类胃癌致病因子,其毒力因子(*cagA*/*vacA*)通过诱导慢性活动性胃炎→萎缩→肠化生→异型增生→腺癌的Correa级联反应驱动胃癌发生。** 在评估Hp毒力因子与胃癌的关联时,需严格控制的混杂因素包括:宿主遗传多态性(IL-1β等)、饮食因素(高盐/腌制食品)、吸烟、年龄、社会经济水平及胃癌家族史。
---
### 一、Hp毒力因子与胃癌的流行病学证据
#### 1. 核心毒力因子及其致病机制
| 毒力因子 | 特征 | 致病机制 |
|---------|------|---------|
| ***cagA***(细胞毒素相关基因A) | 仅存在于部分菌株的*cag*致病岛(*cag* PAI),通过Ⅳ型分泌系统(T4SS)转运CagA毒素至胃黏膜细胞 | 干扰细胞信号通路,促进炎症反应(IL-8、TNF-α释放),被认为是Hp致癌的主要毒力基因[6] |
| ***vacA***(空泡细胞毒素基因A) | 所有菌株均携带,但基因序列高度多态性;功能性VacA毒素导致细胞质空泡化改变 | 基因变异与疾病严重程度相关[6] |
| **尿素酶** | 将尿素水解为二氧化碳和氨,中和胃酸 | 使Hp在胃内酸性环境中存活,对感染胃黏膜至关重要[6] |
| **鞭毛** | 帮助Hp穿过胃黏液层 | 快速到达胃黏膜定植[6] |
#### 2. 流行病学数据
- **全球负担**:Hp感染赋予个体终生患胃癌风险为**1.5–2.0%**,全球每年胃癌负担超过100万例,致死率极高[14][17]。
- **地域分布**:东亚(日本、韩国、中国东部)患病率最高,全球**40%的胃癌病例发生在中国**[14][17]。
**以下为全球Hp感染流行率分布图:**
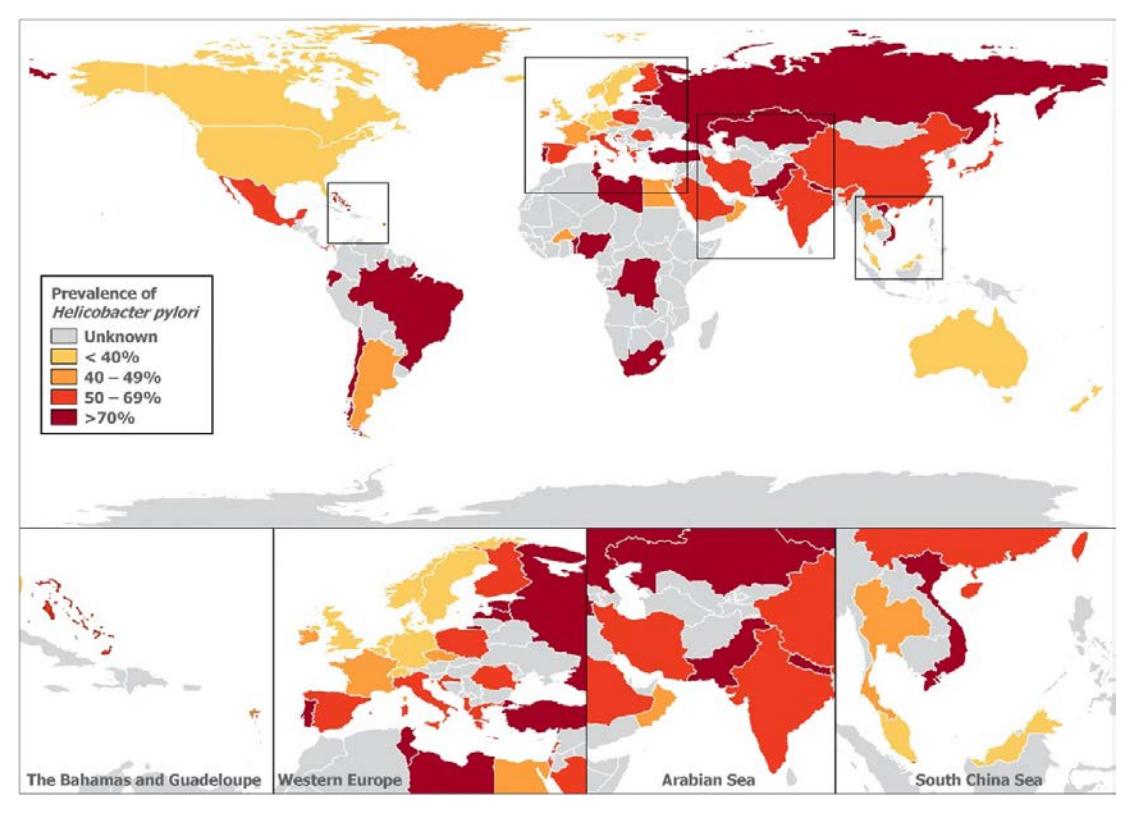
*图:全球幽门螺杆菌感染流行率分布图,显示南美洲、非洲及亚洲部分地区流行率>70%,北美及西欧流行率<40%[Figure 1]*
- **Hp与胃癌的关联强度**:一项长达15年的随访研究显示,Hp感染引发胃癌的**HR为5.8(95% CI: 2.7–15.3)**,而萎缩性胃炎导致胃癌的**HR达9.1(95% CI: 2.9–30.0)**[9]。
- **毒力因子分层**:分子流行病学调查显示,*cagA*阳性菌株感染显著增加胃癌发病风险[6]。
---
### 二、Correa级联反应:从Hp感染到胃癌的演进路径
**以下为胃黏膜从正常组织经癌前阶段进展至早期腺癌的内镜及组织病理学演进图:**
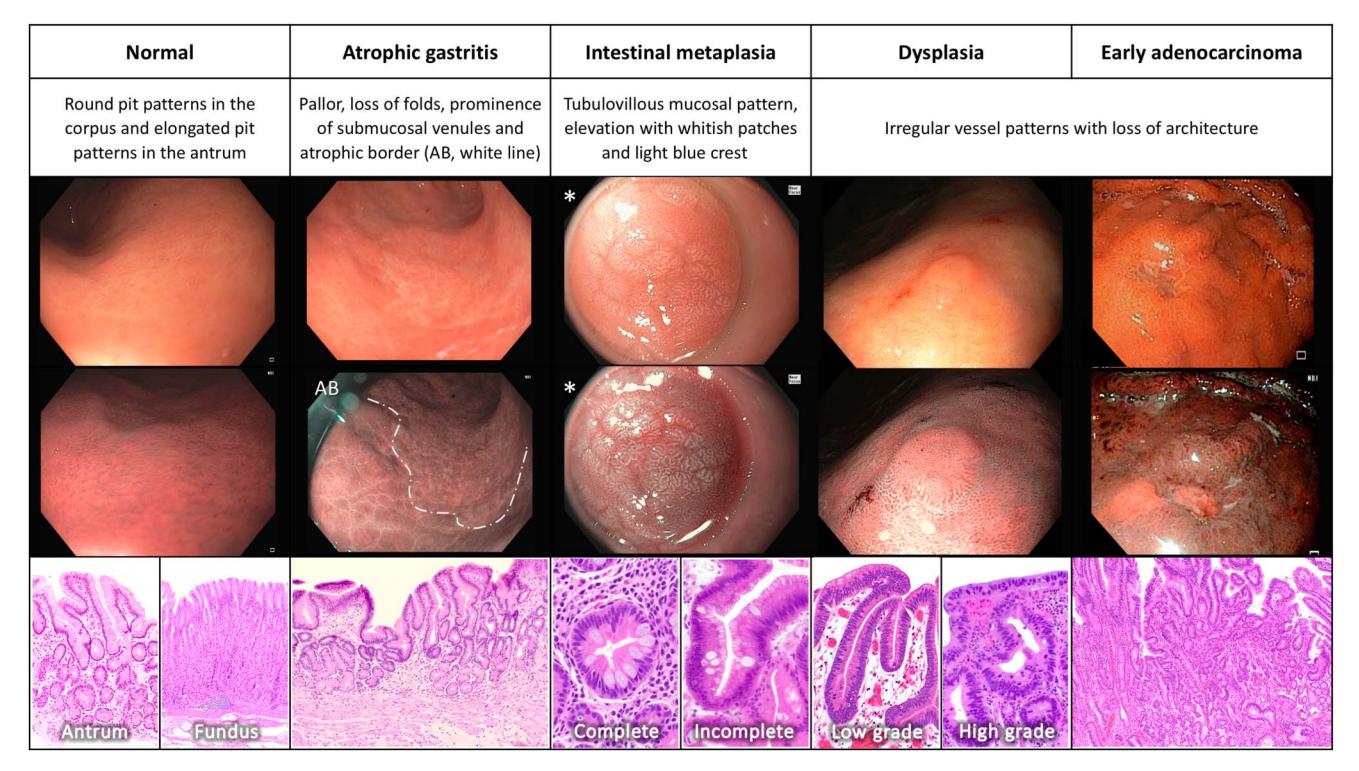
*图:胃黏膜从正常→萎缩性胃炎→肠化生→异型增生→早期腺癌的Correa级联反应内镜及组织病理学表现[Figure 6]*
#### 关键演进节点
| 阶段 | 内镜特征 | 组织病理学特征 | 年胃癌发生率 |
|------|---------|--------------|------------|
| **正常胃黏膜** | 胃体圆形隐窝、胃窦椭圆形隐窝 | 正常腺体结构 | — |
| **萎缩性胃炎** | 黏膜苍白、皱襞消失、黏膜下静脉显露 | 腺体减少/消失 | **0.1%**[9] |
| **肠化生** | 管状绒毛状黏膜模式、白色斑块、"淡蓝嵴"征 | 杯状细胞、Paneth细胞出现 | **0.25%**[9] |
| **异型增生** | 微血管模式不规则、黏膜结构丧失 | 低级别/高级别异型增生 | 显著升高 |
| **早期腺癌** | 黏膜结构完全丧失 | 恶性细胞浸润 | — |
#### 风险分层
- **高风险萎缩性胃炎**:包括重度萎缩、胃窦和胃体部同时肠化生、OLGA/OLGIM Ⅲ/Ⅳ期[16]。
- **广泛萎缩和肠化生**:5年胃癌累积发病率分别为**1.9%–10%**和**5.3%–9.8%**[16]。
- **胃体部病变**:胃体部萎缩、肠化生的病变进展比例较胃窦部高,提示胃体部癌前病变具有更高的病变进展风险[9]。
---
### 三、Hp根除治疗对胃癌的预防作用
#### 核心证据
| 研究/指南 | 关键发现 | 推荐级别 |
|----------|---------|---------|
| 山东临朐临床试验(随访14.7年) | Hp根除治疗可显著降低胃癌发病率,即使对已出现肠化生甚至异型增生者亦有预防作用[8] | — |
| 韩国RCT(NEJM发表) | 对有一级亲属胃癌家族史的Hp阳性患者进行根除治疗可明显降低胃癌发生风险[8] | — |
| 2025 ESGE/EHMSG/ESP指南(MAPS III) | Hp根除降低胃癌风险,在无基线癌前病变者中更明显(**HR 0.37, 95% CI: 0.15–0.95**)[3] | **强推荐/高质量** |
| 2021 Maastricht VI共识 | Hp根除可消除非萎缩性胃炎的活动性炎症,阻止进展为萎缩和肠化生[15] | **A1级** |
| 2021 Maastricht VI共识 | Hp根除可在部分患者中逆转胃萎缩及一定程度逆转肠化生[15] | **A1级** |
#### 根除治疗的时机效应
- **最佳时机**:在发生严重慢性萎缩性胃炎之前根除Hp,胃癌预防效果最显著[15]。
- **成年后任何年龄**:Hp根除在任何成年年龄均可提供胃癌预防机会,但获益幅度随年龄增长而递减[15]。
- **已发生肠化生后**:根除Hp可降低胃癌发生风险,但不能完全消除风险;残留风险与黏膜病变的范围和严重程度及其他宿主风险因素有关[14]。
---
### 四、胃癌风险因素与混杂控制
#### 1. 已确立的风险因素
| 风险因素类别 | 具体因素 | 证据来源 |
|-------------|---------|---------|
| **感染因素** | Hp感染(HR 5.8, 95% CI: 2.7–15.3)[9] | [6][7][9][10][14] |
| **癌前病变** | 萎缩性胃炎(HR 9.1, 95% CI: 2.9–30.0)[9];肠化生(年胃癌发生率0.25%)[9] | [9][16] |
| **遗传因素** | 胃癌一级亲属风险升高2–10倍[9][16];IL-1β基因多态性[14][18] | [9][14][16][18] |
| **饮食因素** | 高盐摄入、腌制食品、抗氧化剂含量低的饮食[14][17] | [14][17][18] |
| **生活方式** | 吸烟、长期饮酒[12][14][18] | [12][14][18] |
| **年龄** | 50岁以上为易感人群[15] | [15] |
#### 2. 需严格控制的混杂因素
在评估Hp毒力因子与胃癌的关联时,以下混杂因素必须控制:
| 混杂因素 | 控制理由 | 控制方法 |
|---------|---------|---------|
| **宿主IL-1β基因多态性** | 决定促炎细胞因子表达,与Hp感染后胃癌风险增加相关[14];携带高表达IL-1β基因多态性者若同时感染*cagA*+或*vacA* s1型菌株,胃癌风险分别增加**25倍**或**87倍**[14] | 基因分型后分层分析或多变量回归 |
| **高盐/腌制食品摄入** | 可增强CagA毒性[18];与胃癌风险增加相关[14][17] | 标准化饮食问卷,纳入回归模型 |
| **吸烟** | 宿主方面与胃癌风险增加相关的因素之一[14][17] | 包-年量化,纳入多变量分析 |
| **社会经济水平** | 儿童时期的社会经济水平是Hp感染流行率的主要决定因素[14] | 匹配或调整教育/收入水平 |
| **年龄** | Hp感染流行率随年龄增长而增加(主要为队列效应)[14];50岁以上为胃癌易感人群[15] | 年龄匹配或连续变量调整 |
| **胃癌家族史** | 一级亲属风险升高2–10倍[9][16] | 分层分析或排除家族史阳性者 |
| **Hp毒力因子共感染** | EBV与Hp均为胃癌独立风险因素,共感染是否增加风险尚不确定[15] | 检测EBV感染状态 |
#### 3. 方法学注意事项
- **病例对照研究中的混杂**:若病例组和对照组的招募比例在不同分层(如研究地点)中不一致,即使该因素本身与疾病风险无关,仍可产生混杂[4]。
- **检测方法差异**:血清学检测(IgG抗体)不能区分现症或既往感染[19];呼气试验(检出率95–98%)[19]和胃镜活检(尿素酶试验检出率90–95%)[19]可确认现症感染。
- **"不可逆点"概念**:在Correa级联反应中,一旦肠化生确立,Hp根除对胃癌风险的降低效果可能有限,强调在癌前病变发生前干预的重要性[4]。
---
### 五、临床管理路径
**以下为2025 ESGE/EHMSG/ESP指南(MAPS III)推荐的胃癌前病变管理路径:**

*图:2025 ESGE/EHMSG/ESP指南(MAPS III)推荐的早期胃肿瘤及癌前病变管理路径,涵盖48项推荐[Figure 2]*
#### 关键推荐
| 推荐内容 | 推荐级别 | 证据质量 |
|---------|---------|---------|
| 非萎缩性胃炎及萎缩性胃炎患者应根除Hp以降低胃癌风险[3] | **强推荐** | **高质量** |
| 已确立肠化生的患者应考虑根除Hp[4] | **条件性推荐** | **中等质量** |
| 胃肿瘤内镜或手术治疗后患者应根除Hp[4] | **强推荐** | **中等质量** |
| 不建议基于遗传状态、出生地或种族进行个体化监测策略[4] | **条件性推荐** | **低质量** |
**以下为2025 ACG指南推荐的非异型增生性胃肠化生管理流程:**
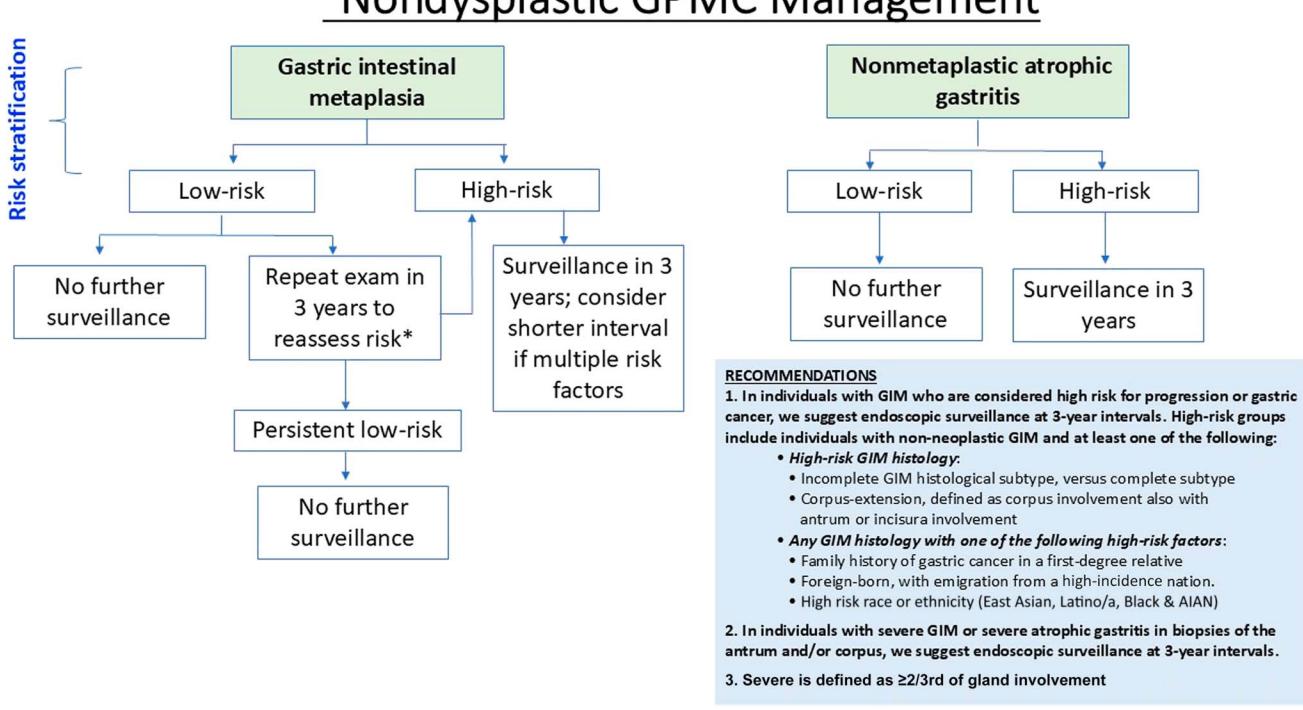
*图:2025 ACG指南推荐的非异型增生性胃肠化生(GIM)风险分层管理流程,低风险者可不监测或3年复查,高风险者每3年监测[Figure 4]*
---
### 六、结论
幽门螺杆菌毒力因子(*cagA*/*vacA*)通过诱导慢性胃炎→萎缩→肠化生→异型增生→腺癌的Corsea级联反应驱动胃癌发生。Hp根除治疗在癌前病变发生前效果最显著(HR 0.37),在已发生肠化生后仍可降低风险但效果有限。在评估Hp毒力因子与胃癌的关联时,需严格控制的混杂因素包括宿主IL-1β基因多态性、高盐/腌制食品摄入、吸烟、年龄、社会经济水平及胃癌家族史。2025年ESGE/EHMSG/ESP指南(MAPS III)及2025年ACG指南均推荐对Hp阳性患者进行根除治疗,并根据萎缩/肠化生的范围和程度进行风险分层监测。
> 本内容基于检索到的文献生成,仅供临床参考。具体诊疗决策应结合患者个体情况并核实原始文献。